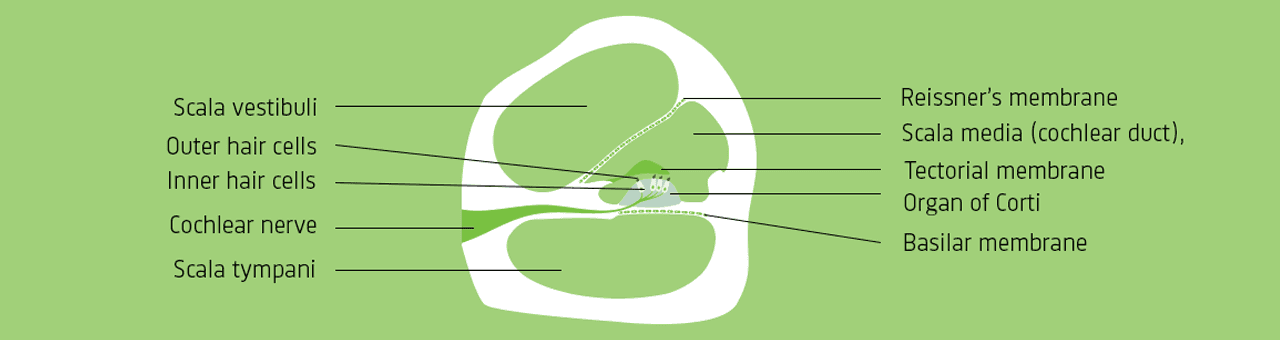
在《人耳》一文的第1部分中,我们通过耳蜗内部的简化示意图(类似于图1中的插图),介绍了基底膜与前庭阶和鼓室阶中的淋巴液之间的相互作用。前庭阶和鼓室阶在基底膜的顶端相连,形成一条很长的连接通道(见图1)。然而,这里仅给出耳蜗简图,常用于重点展示淋巴液与基底膜运动之间的相互作用。
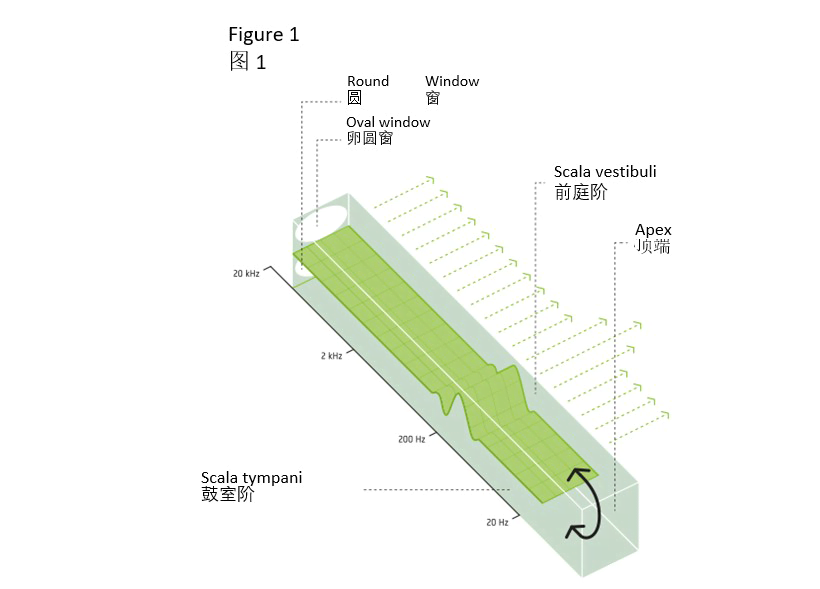
耳蜗横截面详图(见图2)显示了充满淋巴液的第三个腔体——蜗管,蜗管与前庭阶和鼓室阶平行。我们还能看到前庭阶不直接接触基底膜,耳蜗管前庭膜将前庭阶与蜗管分隔开来。
同理,基底膜也将蜗管与鼓室阶分隔开来。前庭阶和鼓室阶中的淋巴液称为外淋巴液。蜗管中的淋巴液则称为内淋巴液。
必须将这两种淋巴液分离开来,以维持内淋巴液中钾离子(K+)的高浓度。
本文中有关说明的依据为当前对转导过程的理解。考虑到难以研究活体耳蜗的内部运作情况,以及已死亡组织中的生命机制不再运行,这方面的研究工作仍有很多不确定性,但这一问题恰好又是当前的研究对象。
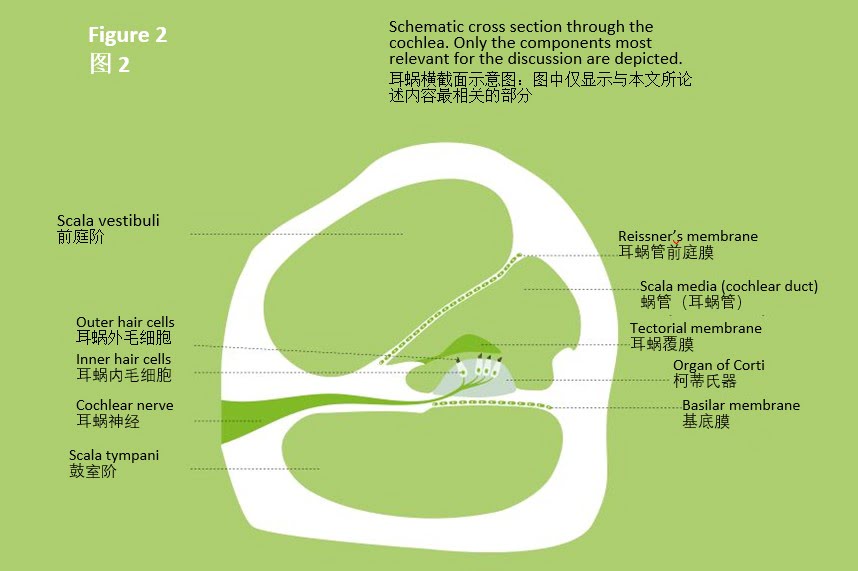
当耳朵在休息状态下,随着钾离子(K+)和钙离子(Ca++)的进入,耳蜗毛在淋巴液与基底膜运动的影响下发生状态变化,耳蜗内毛细胞触发神经脉冲。随后,多余的离子从内毛细胞(见下图中虚线箭头所示部分)中排出,使内毛细胞恢复至默认状态。
基底膜顶部有一个称为“柯蒂氏器”的复杂结构。正是这个实际的转导有机体将运动转化为随后经听觉神经纤维发送至大脑的电化学脉冲。该器官的上层包含成千上万个伸入内淋巴液中的耳蜗内毛细胞。我们在下图中分别标示了耳蜗内毛细胞和耳蜗外毛细胞。
耳蜗内毛细胞
耳蜗内毛细胞将机械运动转化为神经信号(见图3)。可将这些细胞视为微型开关。在闭合状态下,内淋巴液中的钾离子无法进入毛细胞中。然而,基底膜与内淋巴液运动会造成内毛和外毛发生相对偏斜,纤毛尖端细丝打开了离子通道,让钾离子进入。这种浓度变化使另一道门也得以打开,让钙离子(Ca++)进入,从而触发了随后发送至中枢神经系统(CNS)的实际脉冲。接下来,离子迅速从细胞中排出,使毛细胞中的钾离子和钙离子恢复至默认的低浓度状态。
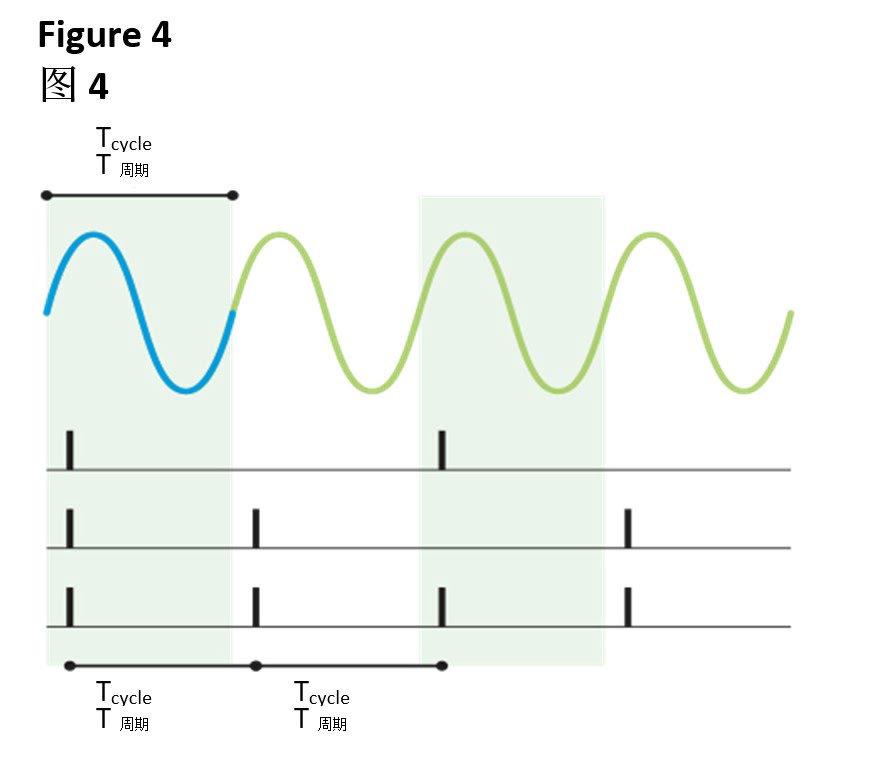
该系列示意图中的第1部分显示了每部分基底膜对某特定频率做出的最强烈反应(见图1)。另外,每部分均与一组特定的听觉神经纤维相关联,并且中枢神经系统会根据发出脉冲的纤维确定相应频率。原则上,一个单脉冲即足以确定某个频率,但实际上并非每个波周期都会出现脉冲(见图4)。相反,每秒产生的脉冲数取决于声强,声音越强,触发的脉冲就越多。
如图4所示,脉冲与声波中的相位信息之间存在锁时性。这种锁相技术在较低频率状态下最精确,随着频率的升高,精度相应降低。听觉系统可借此对比左耳和右耳之间的时序(详见《Waves》杂志第8期中的"Listening in 3D"一文)。由于头部形状以及两耳的间距,左耳或右耳能略早记录来自侧面的信号。除了对比猝发信号的到达时间外,听觉系统还能定向追踪连续不断的声音。
耳蜗外毛细胞
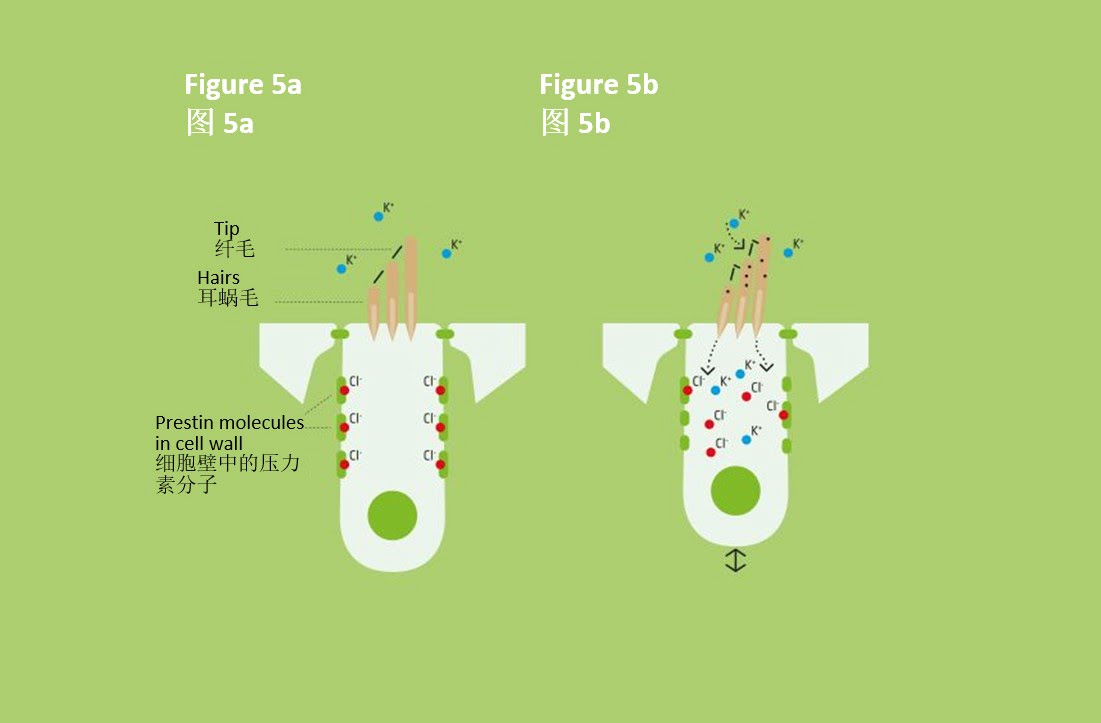
耳蜗外毛细胞不参与转导,但能提高灵敏度。此类细胞的细胞壁上含有一种称为压力素的动力蛋白,在与阴离子(此例中为氯阴离子(Cl–))结合时,压力素的体积会增加。只要毛细胞运动导致钾离子进入,氯阴离子(Cl–)就会从压力素分子中排出,导致压力素体积变小(见图5)。在钾离子浓度再次降低时,阴离子会再次结合,压力素分子恢复至膨胀状态。因此,随着整个细胞的收缩或膨胀,基底膜的振幅相应增大。
外毛细胞的这种电能动性过程对于非常柔和的声音的检测至关重要。研究表明,如果缺少外毛细胞的这一积极作用,我们会丧失20-40dB的听觉灵敏度。如果缺少这种机制,我们就无法听到森林里树叶发出的飒飒作响声、鸟雀轻啼声、图书馆或阅览室环境下的各种寂静之声,或者难以检测到此类噪音。
脉冲锁相原理:两个连续脉冲之间的时间针对同一音调下的一个完整的波周期(或多个波周期),因此总是在同一波周期的相同相位下触发脉冲,此例中,开始后不久即触发脉冲。
外毛细胞的工作原理:细胞内的钾离子(K+)浓度低,当钾离子大量进入后,氯阴离子(Cl–)就会与压力素分子相结合,然后从压力素分子中排出氯阴离子。
听力保护
为何本文用三页篇幅介绍电化学过程?这是为了说明我们的内耳是一个需要呵护的脆弱器官。出生时,人耳大约含有3500个内毛细胞和12000个外毛细胞。不幸的是此类细胞无法再生,一旦此类细胞受损,就会造成永久性失聪。尽管年龄和疾病可能是导致听力损失的两大因素,但导致听力损失(以及很大程度上能避免的听力损失)的一个主要原因就是过度接触噪声。因此,请务必防止耳朵在工作时和闲暇时接触巨响声和过量噪声。